In animal models, apoE is induced within hours to weeks after TBI followed by increased neuronal uptake of apoE-containing lipoproteins through the low-density lipoprotein receptor. In mice, apoE deficiency compromises recovery from acute neurological injuries, including TBI. ApoE also regulates b-amyloid metabolism, a function that underlies its genetic association with Alzheimer��s disease. AD is defined neuropathologically by the presence of extracellular amyloid plaques composed of aggregated Ab peptides and intracellular neurofibrillary tangles consisting of hyperphosphorylated tau. The human apoE4 allele increases AD risk and hastens its onset, whereas apoE2 delays onset and reduces AD risk. Recent microdialysis studies demonstrated that apoE genotype modulates the Ab half-life in brain interstitial fluid, with the 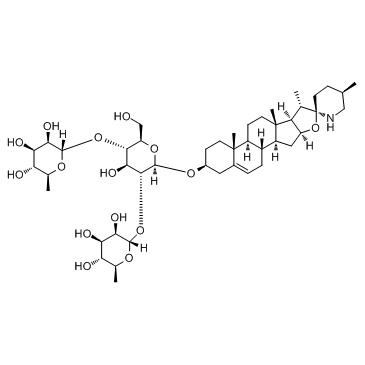 rate of Ab Folinic acid calcium salt pentahydrate degradation accelerated by apoE2 and prolonged by apoE4 compared to apoE3. Although AD risk and age of onset are indisputably modified by apoE genotype, the relationship between apoE genotype and TBI outcome is complex. Some studies report that apoE4 carriers have significantly poorer outcomes compared to non-carriers, while others find no association. Many variables pose challenges to resolving these uncertainties, including Butenafine hydrochloride relatively short follow-up times after TBI and the diversity of subjects with respect to the age, gender, and injury severity. Despite these challenges, a meta-analysis of 14 studies with 2,527 subjects concluded that apoE4 does not affect initial injury severity but rather may compromise recovery at 6 months post-injury. Several epidemiological studies suggest that TBI may increase the risk of dementia, particularly AD, although this association is not always observed. TBI has also been associated with an earlier onset of AD. Both NFTs and amyloid plaques are found in post-mortem TBI brain tissue. However, the widespread white matter involvement in TBI that is not present in AD results in noteworthy differences in the pattern and distribution of their common neuropathological features. For example, TBI neuropathology is largely that of tau deposition, as only approximately 30% of TBI patients also contain Ab deposits. Ab plaques can appear within hours and remain detectable up to 47 years after a single moderatesevere TBI. NFTs are also detectable after a single TBI and are remarkably prominent in mild, repetitive, concussive injuries. Despite the low prevalence of amyloid deposits in the post-mortem TBI brain, amyloid precursor protein accumulation in damaged axons is a striking histological hallmark of diffuse axonal injury. Increased APP in the post-TBI brain is hypothesized to trigger a burst of Ab production that can deposit in amyloid plaques. Intriguingly, microdialysis experiments in brain-injured humans have demonstrated that ISF Ab levels correlate positively with the patient��s Glasgow Coma Score, suggesting that Ab release is associated with recovery of synaptic function, as has been demonstrated in animals. Improving apoE function may therefore facilitate both neuronal repair and Ab clearance after TBI, thus potentially offering both acute and long-term benefits. One method to enhance apoE function is to promote its lipidation by ATP-binding cassette transporter A1, which is the rate-limiting step in generating apoE-containing lipoprotein particles in the central nervous system. ABCA1 deficiency leads to poorly-lipidated apoE in the CNS, and increases amyloid load in AD mice.
rate of Ab Folinic acid calcium salt pentahydrate degradation accelerated by apoE2 and prolonged by apoE4 compared to apoE3. Although AD risk and age of onset are indisputably modified by apoE genotype, the relationship between apoE genotype and TBI outcome is complex. Some studies report that apoE4 carriers have significantly poorer outcomes compared to non-carriers, while others find no association. Many variables pose challenges to resolving these uncertainties, including Butenafine hydrochloride relatively short follow-up times after TBI and the diversity of subjects with respect to the age, gender, and injury severity. Despite these challenges, a meta-analysis of 14 studies with 2,527 subjects concluded that apoE4 does not affect initial injury severity but rather may compromise recovery at 6 months post-injury. Several epidemiological studies suggest that TBI may increase the risk of dementia, particularly AD, although this association is not always observed. TBI has also been associated with an earlier onset of AD. Both NFTs and amyloid plaques are found in post-mortem TBI brain tissue. However, the widespread white matter involvement in TBI that is not present in AD results in noteworthy differences in the pattern and distribution of their common neuropathological features. For example, TBI neuropathology is largely that of tau deposition, as only approximately 30% of TBI patients also contain Ab deposits. Ab plaques can appear within hours and remain detectable up to 47 years after a single moderatesevere TBI. NFTs are also detectable after a single TBI and are remarkably prominent in mild, repetitive, concussive injuries. Despite the low prevalence of amyloid deposits in the post-mortem TBI brain, amyloid precursor protein accumulation in damaged axons is a striking histological hallmark of diffuse axonal injury. Increased APP in the post-TBI brain is hypothesized to trigger a burst of Ab production that can deposit in amyloid plaques. Intriguingly, microdialysis experiments in brain-injured humans have demonstrated that ISF Ab levels correlate positively with the patient��s Glasgow Coma Score, suggesting that Ab release is associated with recovery of synaptic function, as has been demonstrated in animals. Improving apoE function may therefore facilitate both neuronal repair and Ab clearance after TBI, thus potentially offering both acute and long-term benefits. One method to enhance apoE function is to promote its lipidation by ATP-binding cassette transporter A1, which is the rate-limiting step in generating apoE-containing lipoprotein particles in the central nervous system. ABCA1 deficiency leads to poorly-lipidated apoE in the CNS, and increases amyloid load in AD mice.
Respectively reflecting the pronounced lipid mobilization and uptake of CSF lipoproteins
Leave a reply
 detected when the putative N-terminal phosphorylation sites S11 and S38 within the first 40aa of Pex11pb were altered generating putative phospho-mimicking “on” and “off” mutants. Although phosphorylation at other putative sites within Pex11pb is possible, we currently do not have experimental evidence that Pex11pb is
detected when the putative N-terminal phosphorylation sites S11 and S38 within the first 40aa of Pex11pb were altered generating putative phospho-mimicking “on” and “off” mutants. Although phosphorylation at other putative sites within Pex11pb is possible, we currently do not have experimental evidence that Pex11pb is  cytosol. The intra-peroxisomal region between the two transmembrane domains is facing the peroxisomal matrix. We cannot rigorously exclude that parts of this region may interact with the matrix site of the peroxisomal membrane, or are partially buried within the membrane. However, deletion of a glycine-rich stretch within the intra-peroxisomal region did not alter the properties of Pex11pb to promote membrane elongation and division of peroxisomes indicating that parts of its internal region are dispensable for these functions. Based on our results on Pex11pb topology, we analyzed putative functional motifs in its sequence and examined their importance for the membrane-shaping properties of Pex11pb. We focused on the cytosolic N-terminal part of the protein which contains three putative amphipathic helices. Pex11pb was the first peroxisomal membrane protein reported to exhibit a special distribution within the peroxisomal membrane as it was found to concentrate in constriction sites on elongated peroxisomes. Furthermore, it preferentially localized to tubular membrane extensions within pre-peroxisomal membrane compartments. A clear preference for tubular membrane structures was also confirmed in this study, as Pex11pb-Myc accumulated in tubular membrane protrusions extending from enlarged peroxisomes which formed under lipidfree culture conditions. Whereas these observations further support a role for Pex11pb in membrane deformation and elongation, the mechanism of its targeting to and retention within these membrane domains remained unclear. In line with this, it has very recently been reported that DHAcontaining phospholipids directly influence homo-oligomerization of Pex11pb, and that incubation of acyl CoA-oxidase deficient fibroblasts with DHA resulted in hyper-oligomerization of Pex11pb giving rise to high molecular mass complexes ranging from 230�C430 kDa. Intriguingly, these findings imply that Pex11pb action on membrane elongation and thus peroxisome division is modulated by phospholipids within the peroxisomal membrane, which in turn are influenced by peroxisomal lipid metabolism such as fatty acid b-oxidation. In previous
cytosol. The intra-peroxisomal region between the two transmembrane domains is facing the peroxisomal matrix. We cannot rigorously exclude that parts of this region may interact with the matrix site of the peroxisomal membrane, or are partially buried within the membrane. However, deletion of a glycine-rich stretch within the intra-peroxisomal region did not alter the properties of Pex11pb to promote membrane elongation and division of peroxisomes indicating that parts of its internal region are dispensable for these functions. Based on our results on Pex11pb topology, we analyzed putative functional motifs in its sequence and examined their importance for the membrane-shaping properties of Pex11pb. We focused on the cytosolic N-terminal part of the protein which contains three putative amphipathic helices. Pex11pb was the first peroxisomal membrane protein reported to exhibit a special distribution within the peroxisomal membrane as it was found to concentrate in constriction sites on elongated peroxisomes. Furthermore, it preferentially localized to tubular membrane extensions within pre-peroxisomal membrane compartments. A clear preference for tubular membrane structures was also confirmed in this study, as Pex11pb-Myc accumulated in tubular membrane protrusions extending from enlarged peroxisomes which formed under lipidfree culture conditions. Whereas these observations further support a role for Pex11pb in membrane deformation and elongation, the mechanism of its targeting to and retention within these membrane domains remained unclear. In line with this, it has very recently been reported that DHAcontaining phospholipids directly influence homo-oligomerization of Pex11pb, and that incubation of acyl CoA-oxidase deficient fibroblasts with DHA resulted in hyper-oligomerization of Pex11pb giving rise to high molecular mass complexes ranging from 230�C430 kDa. Intriguingly, these findings imply that Pex11pb action on membrane elongation and thus peroxisome division is modulated by phospholipids within the peroxisomal membrane, which in turn are influenced by peroxisomal lipid metabolism such as fatty acid b-oxidation. In previous  identity was not defined. Thus, it is not clear to what extent globally hypomethylated cells are able to commit to and progress along definitive embryonic lineages. The role of DNA methylation in controlling transcription programs during mammalian development and lineage specification has been mainly inferred from genomic methylcytosine profiles in a limited selection of cell types and developmental stages, while little information is available about how DNA methylation and Dnmts actually affect transcription during differentiation. In
identity was not defined. Thus, it is not clear to what extent globally hypomethylated cells are able to commit to and progress along definitive embryonic lineages. The role of DNA methylation in controlling transcription programs during mammalian development and lineage specification has been mainly inferred from genomic methylcytosine profiles in a limited selection of cell types and developmental stages, while little information is available about how DNA methylation and Dnmts actually affect transcription during differentiation. In  bud fate. To determine the earliest time point for the transcriptome analysis, we first analyzed changes in bud morphology during its development, and changes in the expression of key flowering control genes.
bud fate. To determine the earliest time point for the transcriptome analysis, we first analyzed changes in bud morphology during its development, and changes in the expression of key flowering control genes.