Our data suggests that expression-profiling of posttreatment samples could be a possible alternative approach. Some studies have suggested that tumors which develop chemoresistance may acquire certain properties inherent to stem cells, and that chemotherapy treatment leads to a concomitant enrichment of cancer stem cells in vitro. We further demonstrate that the acquired resistance signature is enriched for genes previously identified in embryonic stem cell expression signatures, further suggesting that for gastric cancer, chemoresistance arises from selection of pre-existing cells with particular stem cell characteristics. These acquired resistance signatures were then compared with the intrinsic drug resistance signature of a separate group of 101 non-rebiopsied patients, using gene set comparison analysis of BRBArrayTools. Briefly, this algorithm computed a P-value for each of 2,446 genes to correlate the expression level vs. TTP of these 101 patients using a proportional hazards model. Then it computed mean negative natural logarithm of the P-values of the single gene univariate tests and the proportion of random sets of 2,446 genes with smaller average summary statistics than the LS summaries computed for the real data. The same analysis was 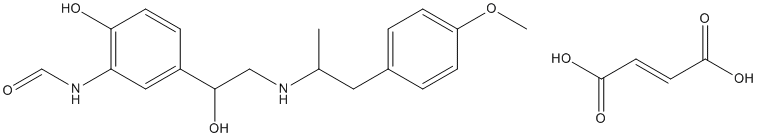 repeated for 633 genes selected at P,0.01. Consistent with results of the hierarchical clustering analyses, the acquired resistance signatures were found to be highly enriched in the ”intrinsic resistance signature” of a separate group of 101 CF-treated patients. LS re-sampling P values were,1025 for both user-defined gene sets selected with different cutoffs to define the acquired resistance signature. Genes overlapping between acquired and intrinsic resistance signatures are listed in Table 2. Figure 2Cb graphically displays that 468 genes upregulated at the chemoresistant state of 22 rebiopsied patients show the concordant overexpression in non-rebiopsied patients with shorter TTP, while 165 genes downregulated at the chemoresistant state show the concordant overexpression in patients with longer TTP. A major finding of this study is the identification of a gene signature that Cinoxacin emerged in association with tumor resistance to CF therapy in patients who initially benefited from CF therapy. Prior genomic predictors for the chemotherapy response, which were developed using pretreatment tissue samples, have demonstrated a mixed performance. Here we demonstrate that the posttreatment samples collected at the time of acquired resistance, although difficult to obtain clinically, contain unique genomic information that can be used to predict the initial response to cytotoxic chemotherapy. No prior studies have explored acquired resistance using Benzoylaconine genome-wide analysis of clinical samples, although 2 prior studies evaluated the gene expression pattern in residual disease after the completion of neoadjuvant chemotherapy. Lee, et al. demonstrated that postchemotherapy tumor gene signatures outperforms baseline signatures and clinical predictors in predicting for pathological response and progression-free survival, although these investigators collected posttreatment breast tumors 3 weeks after chemotherapy, not at the time of progressive disease as in our study. Our data is consistent with the aforementioned study that comparing postchemotherapy and prechemotherapy gene expression signatures might be a feasible approach to the identification of predictive signatures. Also, our data provides the first genomic evidence in clinical samples supporting a conventional model for the emergence of acquired resistance whereby resistance emerges through a selective, clonal outgrowth of small populations of pre-existing, chemoresistant tumor cells. While the ”72-gene acquired resistance signature” was developed mainly for potential clinical utility, it contains several overexpressed genes that have been shown to lead to chemoresistance. TRAP1 overexpression leads to 5-fluorouracil-, oxaliplatin- and irinotecanresistant phenotypes in different neoplastic cells.
repeated for 633 genes selected at P,0.01. Consistent with results of the hierarchical clustering analyses, the acquired resistance signatures were found to be highly enriched in the ”intrinsic resistance signature” of a separate group of 101 CF-treated patients. LS re-sampling P values were,1025 for both user-defined gene sets selected with different cutoffs to define the acquired resistance signature. Genes overlapping between acquired and intrinsic resistance signatures are listed in Table 2. Figure 2Cb graphically displays that 468 genes upregulated at the chemoresistant state of 22 rebiopsied patients show the concordant overexpression in non-rebiopsied patients with shorter TTP, while 165 genes downregulated at the chemoresistant state show the concordant overexpression in patients with longer TTP. A major finding of this study is the identification of a gene signature that Cinoxacin emerged in association with tumor resistance to CF therapy in patients who initially benefited from CF therapy. Prior genomic predictors for the chemotherapy response, which were developed using pretreatment tissue samples, have demonstrated a mixed performance. Here we demonstrate that the posttreatment samples collected at the time of acquired resistance, although difficult to obtain clinically, contain unique genomic information that can be used to predict the initial response to cytotoxic chemotherapy. No prior studies have explored acquired resistance using Benzoylaconine genome-wide analysis of clinical samples, although 2 prior studies evaluated the gene expression pattern in residual disease after the completion of neoadjuvant chemotherapy. Lee, et al. demonstrated that postchemotherapy tumor gene signatures outperforms baseline signatures and clinical predictors in predicting for pathological response and progression-free survival, although these investigators collected posttreatment breast tumors 3 weeks after chemotherapy, not at the time of progressive disease as in our study. Our data is consistent with the aforementioned study that comparing postchemotherapy and prechemotherapy gene expression signatures might be a feasible approach to the identification of predictive signatures. Also, our data provides the first genomic evidence in clinical samples supporting a conventional model for the emergence of acquired resistance whereby resistance emerges through a selective, clonal outgrowth of small populations of pre-existing, chemoresistant tumor cells. While the ”72-gene acquired resistance signature” was developed mainly for potential clinical utility, it contains several overexpressed genes that have been shown to lead to chemoresistance. TRAP1 overexpression leads to 5-fluorouracil-, oxaliplatin- and irinotecanresistant phenotypes in different neoplastic cells.
Due to the difficulty of identifying robust gene signatures in tumors from populations with large genetic variation
Leave a reply
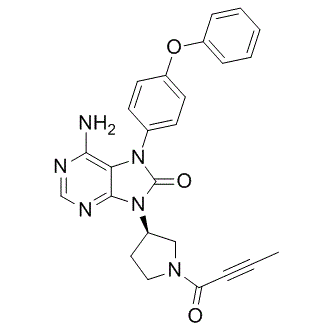 The endoplasmic reticulum is the folding and assembly site for resident structural proteins and enzymes, as well as for secretory and plasma
The endoplasmic reticulum is the folding and assembly site for resident structural proteins and enzymes, as well as for secretory and plasma 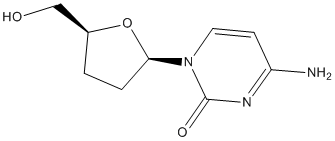 of its long history as an informative metric of local adaptation. We first used the five demographic models inferred from the pairwise interpopulation comparisons to generate neutral distributions of expected FST for silent sites and for all sites. For each locus and model, we calculated FST from 10,000 single locus coalescent simulations drawn from our estimated posterior distributions. All simulations used the relevant length and observed hw for each locus. We present here two significant advances towards understanding
of its long history as an informative metric of local adaptation. We first used the five demographic models inferred from the pairwise interpopulation comparisons to generate neutral distributions of expected FST for silent sites and for all sites. For each locus and model, we calculated FST from 10,000 single locus coalescent simulations drawn from our estimated posterior distributions. All simulations used the relevant length and observed hw for each locus. We present here two significant advances towards understanding 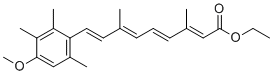 micrometastases was not biased by intravascular delivery of tumor cells, we characterized a syngeneic model of spontaneous brain metastasis. Brain sections were examined for spontaneous micrometastases 5�C7 weeks after orthotopic injection of 4T1-GFP cells into the mammary fat pad. The growth and morphological characteristics of these colonies were indistinguishable from those derived from intracardiac delivery of cells demonstrating both intact GLUT-1positive vasculature and angiotropic spread upon adjacent capillaries. To assess the clinical relevance of frequent vascular cooption in experimental metastasis, we asked whether tumor cells within human brain metastasis specimens displayed a similar vascular association. In brain micrometastases and tumors with carcinomatous CNS spread that could be considered at the early stages of parenchymal colonization, the patterns were similar to those in the experimental models above, with the tumor cells in these brain metastases appearing to track along the blood vessels. Quantitation of vascular cooption in these cases from primary tumors of
micrometastases was not biased by intravascular delivery of tumor cells, we characterized a syngeneic model of spontaneous brain metastasis. Brain sections were examined for spontaneous micrometastases 5�C7 weeks after orthotopic injection of 4T1-GFP cells into the mammary fat pad. The growth and morphological characteristics of these colonies were indistinguishable from those derived from intracardiac delivery of cells demonstrating both intact GLUT-1positive vasculature and angiotropic spread upon adjacent capillaries. To assess the clinical relevance of frequent vascular cooption in experimental metastasis, we asked whether tumor cells within human brain metastasis specimens displayed a similar vascular association. In brain micrometastases and tumors with carcinomatous CNS spread that could be considered at the early stages of parenchymal colonization, the patterns were similar to those in the experimental models above, with the tumor cells in these brain metastases appearing to track along the blood vessels. Quantitation of vascular cooption in these cases from primary tumors of  Figure 1A, GFP-labeled mutant httex1, containing an expanded polyglutamine repeat associated with juvenile HD onset, rapidly formed cytoplasmic and perinuclear aggregates in ST14A cells after 48 hours, whereas httex1 containing a non-amyloidogenic polyglutamine repeat remained completely diffuse.
Figure 1A, GFP-labeled mutant httex1, containing an expanded polyglutamine repeat associated with juvenile HD onset, rapidly formed cytoplasmic and perinuclear aggregates in ST14A cells after 48 hours, whereas httex1 containing a non-amyloidogenic polyglutamine repeat remained completely diffuse.