Survival and proliferation of bacteria depend on their expressing the right amounts of the right genes at the right time. However, what is “right” at any given time-point will vary with the environmental conditions and the specific growth phase. Bacteria often respond to these changing environmental stimuli by switching the expression of specific genes “on” or “off”. To ensure that target gene expression is optimal, will require finetuning of the regulatory parameters that control the switch, and this fine-tuning can affect each individual step of gene expression. In bacteria, gene expression is frequently controlled by proteins that activate or repress transcription by binding to specific DNA sequences close to a promoter. The DNA binding activity of these transcription factors is triggered by small molecules or, less often, by protein-protein interactions. Tet repressor is a paradigm for a bacterial transcription factor that responds directly to an environmental signal by binding a small molecule. TetR regulates transcription of the resistance protein TetA in at least 14 different efflux-type tetracycline resistance determinants found predominantly in Gram-negative bacteria. Repression by TetR has to be tight, because overproduction or constitutive expression of the membrane transporter TetA strongly reduces bacterial fitness. But, at the same time, induction must be sensitive to ensure that TetA is translated before the antibiotic reaches an intracellular level that inhibits translation. Although seemingly conflicting, these requirements are met by the exceptionally high specificity of TetR for its cognate binding site tetO over non-specific DNA and by its unusually high affinity for tetracyclines. Such favorable properties have made TetR a very popular tool for many different applications, including conditional gene expression in both pro- and eukaryotes, overexpression of heterologous proteins or artificial genetic circuits in synthetic biology with highly diverse architectures. So far, all applications using Tet regulation have relied on tetracycline or its analogs as inducers. The recent discovery that peptides can also specifically induce TetR when they are fused to a carrier protein added a new quality to Tet regulation. These inducing peptides, called TIP, bind to the tetracycline-binding pocket of TetR and elicit an allosteric conformational Lomitapide Mesylate change that leads to the complete loss of DNAbinding activity. This turned TetR from an exclusively small-molecule-controlled protein into a downstream effector in a protein signal transduction pathway. Examples of protein-induced regulation of gene expression are not so common in bacteria, but have been found among the major transcription factor families. Information transfer by protein-mediated signal transduction not only introduces new ways to manipulate TetR-based genetic networks in synthetic biology. It also allows to gather proteomic data by determining protein expression profiles after tagging many different proteins with TIP and monitoring their expression by genetic LOUREIRIN-B readout of the TetR-controlled reporter gene. Compared with the intensely studied and well-characterized induction of gene expression by tetracyclines, the 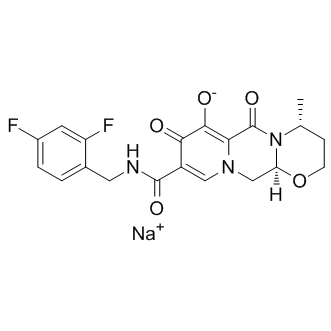 parameters for sensitive and efficient control of a Tet-regulated reporter gene by a protein-based inducer are still largely unknown. While the basic functionality of TIP-mediated induction of TetR has been demonstrated in Escherichia coli and in Staphylococcus aureus, major obstacles must still be overcome, if this system is to be used effectively in more sophisticated applications, like those mentioned above. In the examples published so far, efficient induction of TetR was only achieved after strong overproduction of the TIP-containing fusion protein from a multicopy plasmid.
parameters for sensitive and efficient control of a Tet-regulated reporter gene by a protein-based inducer are still largely unknown. While the basic functionality of TIP-mediated induction of TetR has been demonstrated in Escherichia coli and in Staphylococcus aureus, major obstacles must still be overcome, if this system is to be used effectively in more sophisticated applications, like those mentioned above. In the examples published so far, efficient induction of TetR was only achieved after strong overproduction of the TIP-containing fusion protein from a multicopy plasmid.
The TIP coding sequence was fused to a gene at its native locus in the genome induction of TetR
Leave a reply
 response or visual discrimination. Thus these findings are consistent with the idea that CIE exposure reduces the ability of the mPFC to flexibly modulate behavior during changing environmental situations that require increased attentional control. Medial PFC networks control executive functions and behavioral flexibility, and in humans they may regulate cognitive control over alcohol intake. Thus the CIEinduced changes in glutamatergic transmission in mPFC pyramidal cells that we describe here may contribute to the cognitive impairments and loss of behavioral control seen in alcoholdependent subjects. On the day following turn bias, mice were trained on a response discrimination task. For this task, the mice were required to always turn in the opposite direction of its turn bias to obtain reward. Mice started from one of 3 arms to discourage them from using an allocentric spatial strategy to locate the food.
response or visual discrimination. Thus these findings are consistent with the idea that CIE exposure reduces the ability of the mPFC to flexibly modulate behavior during changing environmental situations that require increased attentional control. Medial PFC networks control executive functions and behavioral flexibility, and in humans they may regulate cognitive control over alcohol intake. Thus the CIEinduced changes in glutamatergic transmission in mPFC pyramidal cells that we describe here may contribute to the cognitive impairments and loss of behavioral control seen in alcoholdependent subjects. On the day following turn bias, mice were trained on a response discrimination task. For this task, the mice were required to always turn in the opposite direction of its turn bias to obtain reward. Mice started from one of 3 arms to discourage them from using an allocentric spatial strategy to locate the food. In addition, by using a conformation specific antibody anti-Ab oligomers, we demonstrated that hNGFP61S/R100E prevents or reduces the accumulation of Ab oligomers, considered the earliest and most synaptotoxic forms of Ab. Thus, hNGFP61S/R100E exerts an anti-amyloidogenic effect in vivo, in a disease-relevant FAD-based model. From a mechanistic point of view, this might involve a generalized neuroprotective activity by the neurotrophin, at multiple levels, by slowing the generation of Ab peptide and Ab oligomers, by reducing the microgliosis and astrogliosis and/or, possibly, by increasing the clearance of Ab peptides, thereby leading to the observed reduced plaque load and Ab oligomer levels. The observed effects on astrocytes and microglia are consistent with both these cell types expressing TrkA and p75NTR NGF receptors, in normal and pathological conditions. Reactive astrocytes, present in APP xPS1 brain due to amyloid pathology, also express higher levels of TrkA in human AD. We found a reduction of both microgliosis and
In addition, by using a conformation specific antibody anti-Ab oligomers, we demonstrated that hNGFP61S/R100E prevents or reduces the accumulation of Ab oligomers, considered the earliest and most synaptotoxic forms of Ab. Thus, hNGFP61S/R100E exerts an anti-amyloidogenic effect in vivo, in a disease-relevant FAD-based model. From a mechanistic point of view, this might involve a generalized neuroprotective activity by the neurotrophin, at multiple levels, by slowing the generation of Ab peptide and Ab oligomers, by reducing the microgliosis and astrogliosis and/or, possibly, by increasing the clearance of Ab peptides, thereby leading to the observed reduced plaque load and Ab oligomer levels. The observed effects on astrocytes and microglia are consistent with both these cell types expressing TrkA and p75NTR NGF receptors, in normal and pathological conditions. Reactive astrocytes, present in APP xPS1 brain due to amyloid pathology, also express higher levels of TrkA in human AD. We found a reduction of both microgliosis and 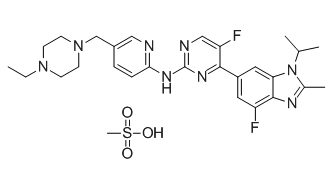 For example, while electrophysiological recordings were restricted to layer V pyramidal neurons in the prelimbic subregion of the mPFC, Western blotting was carried out using tissue punches that included not only all layers of the prelimbic cortex, but also portions of the subregions surrounding the prelimbic PFC. Layer-specific effects on synaptic transmission have been observed for several drugs of abuse including alcohol. Furthermore, function with a return to baseline density so as to allow for subsequent plasticity events.
For example, while electrophysiological recordings were restricted to layer V pyramidal neurons in the prelimbic subregion of the mPFC, Western blotting was carried out using tissue punches that included not only all layers of the prelimbic cortex, but also portions of the subregions surrounding the prelimbic PFC. Layer-specific effects on synaptic transmission have been observed for several drugs of abuse including alcohol. Furthermore, function with a return to baseline density so as to allow for subsequent plasticity events.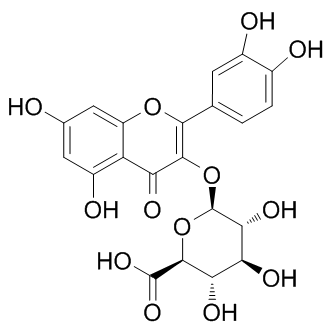 the same binding scores of 0.982. This is supported by the fact that the nonsteroidal anti-inflammatory drugs could produce therapeutic activities through the inhibition of cyclooxygenase. With no exception, for all the remaining interactions, the predicted ligands of certain receptors are invariably similar in structure with those confirmed ligands as mentioned above. All these outcomes enhance the strength of our proposed methods for realistic drugtarget interaction prediction application. Subsequently, a comprehensive network describing the drugtarget interactions was constructed. In order to make it clear and simple, the top 500 scoring drug-target interactions were used to generate a bipartite graph of drug-target interactions for illustrating the complex relationships between drugs and enzymes, in which a compound and a protein are connected to each other if the protein is a predicted target of the compound. Figure 7 shows a global view of this network with color and shape-coded nodes. To explore the topological and global properties of this drug-target network, the centralization, heterogeneity and node degree distribution were analyzed. The centralization and heterogeneity analysis shows the network centralization and heterogeneity degrees are 0.463 and 3.661, respectively, indicating that a few nodes are more central that the other ones in this net, i.e., the drugtarget space is biased toward certain compounds and proteins.
the same binding scores of 0.982. This is supported by the fact that the nonsteroidal anti-inflammatory drugs could produce therapeutic activities through the inhibition of cyclooxygenase. With no exception, for all the remaining interactions, the predicted ligands of certain receptors are invariably similar in structure with those confirmed ligands as mentioned above. All these outcomes enhance the strength of our proposed methods for realistic drugtarget interaction prediction application. Subsequently, a comprehensive network describing the drugtarget interactions was constructed. In order to make it clear and simple, the top 500 scoring drug-target interactions were used to generate a bipartite graph of drug-target interactions for illustrating the complex relationships between drugs and enzymes, in which a compound and a protein are connected to each other if the protein is a predicted target of the compound. Figure 7 shows a global view of this network with color and shape-coded nodes. To explore the topological and global properties of this drug-target network, the centralization, heterogeneity and node degree distribution were analyzed. The centralization and heterogeneity analysis shows the network centralization and heterogeneity degrees are 0.463 and 3.661, respectively, indicating that a few nodes are more central that the other ones in this net, i.e., the drugtarget space is biased toward certain compounds and proteins.