However, various topologies were proposed for Pex11 proteins in different organisms,, and the related Pex11pc was recently reported to dock on the cytosolic site of the peroxisomal membrane. Furthermore, the Tulathromycin B predicted position of the first transmembrane domain within human Pex11pb varies greatly, depending on the in silico search algorithm used, thus resulting e.g. in the designation of Pex11pb as a tail-anchored membrane protein. In the present study, we characterized a newly available Pex11pb antibody directed against an epitope within the putative internal region to determine the topology of Pex11pb. Using differential permeabilization, we confirmed that the epitope recognized by the Pex11pb antibody is only accessible under conditions which permeabilize the peroxisomal membrane. Proteinase K digest of intact peroxisomes and subsequent immunoblotting with the Pex11pb antibody revealed a protease-resistant fragment of approximately 17 kDa, which was degraded upon membrane permeabilization with either Triton X-100 or sonication. The fragment size is consistent with the localization of the first transmembrane domain between aa 90�C110, and the second one between aa 230�C255. These data clearly demonstrate that Pex11pb is an integral membrane protein with two transmembrane spanning domains and N- and C-termini directed towards the 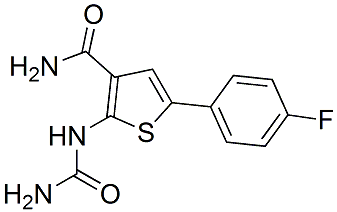 cytosol. The intra-peroxisomal region between the two transmembrane domains is facing the peroxisomal matrix. We cannot rigorously exclude that parts of this region may interact with the matrix site of the peroxisomal membrane, or are partially buried within the membrane. However, deletion of a glycine-rich stretch within the intra-peroxisomal region did not alter the properties of Pex11pb to promote membrane elongation and division of peroxisomes indicating that parts of its internal region are dispensable for these functions. Based on our results on Pex11pb topology, we analyzed putative functional motifs in its sequence and examined their importance for the membrane-shaping properties of Pex11pb. We focused on the cytosolic N-terminal part of the protein which contains three putative amphipathic helices. Pex11pb was the first peroxisomal membrane protein reported to exhibit a special distribution within the peroxisomal membrane as it was found to concentrate in constriction sites on elongated peroxisomes. Furthermore, it preferentially localized to tubular membrane extensions within pre-peroxisomal membrane compartments. A clear preference for tubular membrane structures was also confirmed in this study, as Pex11pb-Myc accumulated in tubular membrane protrusions extending from enlarged peroxisomes which formed under lipidfree culture conditions. Whereas these observations further support a role for Pex11pb in membrane deformation and elongation, the mechanism of its targeting to and retention within these membrane domains remained unclear. In line with this, it has very recently been reported that DHAcontaining phospholipids directly influence homo-oligomerization of Pex11pb, and that incubation of acyl CoA-oxidase deficient fibroblasts with DHA resulted in hyper-oligomerization of Pex11pb giving rise to high molecular mass complexes ranging from 230�C430 kDa. Intriguingly, these findings imply that Pex11pb action on membrane elongation and thus peroxisome division is modulated by phospholipids within the peroxisomal membrane, which in turn are influenced by peroxisomal lipid metabolism such as fatty acid b-oxidation. In previous Butenafine hydrochloride studies, we showed that the addition of polyunsaturated fatty acids promotes the elongatio.
cytosol. The intra-peroxisomal region between the two transmembrane domains is facing the peroxisomal matrix. We cannot rigorously exclude that parts of this region may interact with the matrix site of the peroxisomal membrane, or are partially buried within the membrane. However, deletion of a glycine-rich stretch within the intra-peroxisomal region did not alter the properties of Pex11pb to promote membrane elongation and division of peroxisomes indicating that parts of its internal region are dispensable for these functions. Based on our results on Pex11pb topology, we analyzed putative functional motifs in its sequence and examined their importance for the membrane-shaping properties of Pex11pb. We focused on the cytosolic N-terminal part of the protein which contains three putative amphipathic helices. Pex11pb was the first peroxisomal membrane protein reported to exhibit a special distribution within the peroxisomal membrane as it was found to concentrate in constriction sites on elongated peroxisomes. Furthermore, it preferentially localized to tubular membrane extensions within pre-peroxisomal membrane compartments. A clear preference for tubular membrane structures was also confirmed in this study, as Pex11pb-Myc accumulated in tubular membrane protrusions extending from enlarged peroxisomes which formed under lipidfree culture conditions. Whereas these observations further support a role for Pex11pb in membrane deformation and elongation, the mechanism of its targeting to and retention within these membrane domains remained unclear. In line with this, it has very recently been reported that DHAcontaining phospholipids directly influence homo-oligomerization of Pex11pb, and that incubation of acyl CoA-oxidase deficient fibroblasts with DHA resulted in hyper-oligomerization of Pex11pb giving rise to high molecular mass complexes ranging from 230�C430 kDa. Intriguingly, these findings imply that Pex11pb action on membrane elongation and thus peroxisome division is modulated by phospholipids within the peroxisomal membrane, which in turn are influenced by peroxisomal lipid metabolism such as fatty acid b-oxidation. In previous Butenafine hydrochloride studies, we showed that the addition of polyunsaturated fatty acids promotes the elongatio.
The cytosol suggesting a transmembrane protein with two membrane spanning domains
Leave a reply
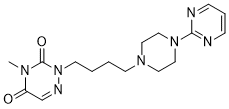 identity was not defined. Thus, it is not clear to what extent globally hypomethylated cells are able to commit to and progress along definitive embryonic lineages. The role of DNA methylation in controlling transcription programs during mammalian development and lineage specification has been mainly inferred from genomic methylcytosine profiles in a limited selection of cell types and developmental stages, while little information is available about how DNA methylation and Dnmts actually affect transcription during differentiation. In
identity was not defined. Thus, it is not clear to what extent globally hypomethylated cells are able to commit to and progress along definitive embryonic lineages. The role of DNA methylation in controlling transcription programs during mammalian development and lineage specification has been mainly inferred from genomic methylcytosine profiles in a limited selection of cell types and developmental stages, while little information is available about how DNA methylation and Dnmts actually affect transcription during differentiation. In 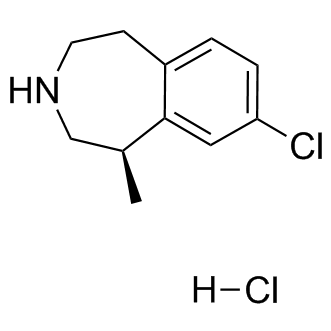 bud fate. To determine the earliest time point for the transcriptome analysis, we first analyzed changes in bud morphology during its development, and changes in the expression of key flowering control genes.
bud fate. To determine the earliest time point for the transcriptome analysis, we first analyzed changes in bud morphology during its development, and changes in the expression of key flowering control genes.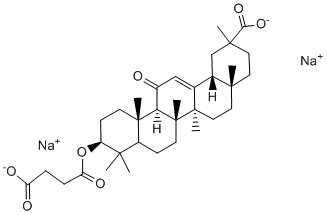 that of HIV-1. From a seed no bigger than that of a cucumber, California’s coastal redwood tree can grow to a height of more than 350 feet. At the same time, aquatic watermeal plants are so small that they resemble specks of cornmeal. The nature of the mechanisms controlling the size and shape of organs is an important but still unanswered question of developmental biology. Two factors determine the size of mature organs: cell number and cell size. A current model for metazoans proposes that cell number and cell size are controlled by distinct proliferation and growth signals that negatively affect each other; when cellular size is increased, cell count is reduced and vice versa. This regulation ensures that induced alterations in cell proliferation are compensated for by changes in cell size, resulting in little net
that of HIV-1. From a seed no bigger than that of a cucumber, California’s coastal redwood tree can grow to a height of more than 350 feet. At the same time, aquatic watermeal plants are so small that they resemble specks of cornmeal. The nature of the mechanisms controlling the size and shape of organs is an important but still unanswered question of developmental biology. Two factors determine the size of mature organs: cell number and cell size. A current model for metazoans proposes that cell number and cell size are controlled by distinct proliferation and growth signals that negatively affect each other; when cellular size is increased, cell count is reduced and vice versa. This regulation ensures that induced alterations in cell proliferation are compensated for by changes in cell size, resulting in little net  this region is important for normal cardiac development. How can we explain these divergent observations? One possibility is that the SRJ is indeed dispensable for mammalian heart development, and that the observations from patients may be misleading. Supporting this idea, there is considerable lack of conservation in this domain between placental and non-placental mammals. Furthermore, the mutations in the SRJ do not significantly affect the disordered nature of the domain, indicating that it may be able to accommodate mutations without adversely affecting the overall structure and function of the protein. Moreover, in no case has it been shown that the mutation has either arisen de novo or been transmitted from an affected parent. Another possibility that partially reconciles the mouse and human observations is that variants found clustered in the SRJ in CHD patients may not, by themselves.
this region is important for normal cardiac development. How can we explain these divergent observations? One possibility is that the SRJ is indeed dispensable for mammalian heart development, and that the observations from patients may be misleading. Supporting this idea, there is considerable lack of conservation in this domain between placental and non-placental mammals. Furthermore, the mutations in the SRJ do not significantly affect the disordered nature of the domain, indicating that it may be able to accommodate mutations without adversely affecting the overall structure and function of the protein. Moreover, in no case has it been shown that the mutation has either arisen de novo or been transmitted from an affected parent. Another possibility that partially reconciles the mouse and human observations is that variants found clustered in the SRJ in CHD patients may not, by themselves.