We suspect dPRL-1/dPRL-1NC may physically interfere with either Src or an effector of Src function. While dPRL-1s ability to inhibit growth is in concordance with one report from the mammalian literature, there are certainly differences to highlight Nutlin-3 between Drosophila and mammalian studies. Sequence analysis shows that the aspartate, that serves as a proton donor is present in Drosophila but not in the context of the WPD loop, as seen in mammalian PRL family members. While this aspartate is also not found in WPD loop in other PTPs like VHR, cdc14, and PTEN, it may point to different substrates between mammals and flies. In addition, catalytic activity of mammalian PRL1 is regulated by the redox environment,,, and thought to exist in an inactive conformation under normal cellular conditions. Possibly, differences in redox AG-013736 319460-85-0 regulation between Drosophila and cultured mammalian cells could account for differing outcomes in response to PRL-1 overexpression. For example, altered redox environments in transformed cells could switch PRLs to an abnormal, catalytically active state. Another important difference between Drosophila and mammals may be the p53 network. While supporting the model that PRL-3 is a transcriptional target of p53, Min et al., report that PRL-3 then functions in 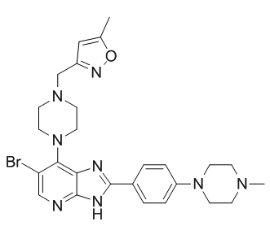 a negative, autoregulatory loop by decreasing levels of p53, which would help transform cells. They identify MDM2 and PIRH2 as the important players in this pathway; but since neither protein is found in Drosophila, this oncogenic path is not conserved. In spite of the differences between mammals and Drosophila, flies have successfully informed numerous mechanisms that contribute to human cancer biology. We have established a new system that has revealed novel characteristics of the PRL family and will help decipher the role PRLs play in cancers. The paradigm of cancer treatment has been dramatically changed by the introduction of small molecular compounds that target the “Achilles’heel” of cancer cells. The proteasome is a proteolytic machinery that executes the degradation of polyubiquitinated proteins to maintain cellular homeostasis. Cancer cells are very sensitive to proteotoxic stress because of intracellular protein overload due to rapid cell cycling and apoptosis inhibition. This feature makes proteasome inhibition a unique and effective way to kill cancer cells that can tolerate conventional therapies. Bortezomib is the first proteasome inhibitor approved for clinical application, which preferentially targets ?1 and ?5 subunits of the proteasome. This drug is particularly effective for multiple myeloma, because it accelerates the unfolded protein response via down-regulation of histone deacetylases and targets cell adhesion-mediated drug resistance via down-regulation of very late antigen-4. Accordingly, bortezomib is now indispensable for the treatment of MM in combination with other anti-cancer drugs including alkylating agents, corticosteroids and HDAC inhibitors. Although bortezomib therapy is a major advance in clinical oncology, there are at least three major problems to be resolved as soon as possible. First, bortezomib has several possible off-target toxicities. Second, the development of intrinsic and acquired resistance to bortezomib is an emerging problem. Third, bortezomib should be administered intravenously on biweekly schedules with treatment periods extending for 6 months or more.
a negative, autoregulatory loop by decreasing levels of p53, which would help transform cells. They identify MDM2 and PIRH2 as the important players in this pathway; but since neither protein is found in Drosophila, this oncogenic path is not conserved. In spite of the differences between mammals and Drosophila, flies have successfully informed numerous mechanisms that contribute to human cancer biology. We have established a new system that has revealed novel characteristics of the PRL family and will help decipher the role PRLs play in cancers. The paradigm of cancer treatment has been dramatically changed by the introduction of small molecular compounds that target the “Achilles’heel” of cancer cells. The proteasome is a proteolytic machinery that executes the degradation of polyubiquitinated proteins to maintain cellular homeostasis. Cancer cells are very sensitive to proteotoxic stress because of intracellular protein overload due to rapid cell cycling and apoptosis inhibition. This feature makes proteasome inhibition a unique and effective way to kill cancer cells that can tolerate conventional therapies. Bortezomib is the first proteasome inhibitor approved for clinical application, which preferentially targets ?1 and ?5 subunits of the proteasome. This drug is particularly effective for multiple myeloma, because it accelerates the unfolded protein response via down-regulation of histone deacetylases and targets cell adhesion-mediated drug resistance via down-regulation of very late antigen-4. Accordingly, bortezomib is now indispensable for the treatment of MM in combination with other anti-cancer drugs including alkylating agents, corticosteroids and HDAC inhibitors. Although bortezomib therapy is a major advance in clinical oncology, there are at least three major problems to be resolved as soon as possible. First, bortezomib has several possible off-target toxicities. Second, the development of intrinsic and acquired resistance to bortezomib is an emerging problem. Third, bortezomib should be administered intravenously on biweekly schedules with treatment periods extending for 6 months or more.
The development of orally bioavailable PIs with distinct mode of action is a possible way to circumvent these issues
Leave a reply